分类:论文范文 发表时间:2021-06-10 10:49
摘要:【目的】对重庆武陵山区10个野生芒居群的17个表型性状进行遗传多样性研究。【方法】对17个表型性状进行方差分析、多样性指数分析、相关性分析、聚类分析及主成分分析等。【结果】10个居群的遗传变异主要体现在居群内,居群内多样性指数为1.88。聚类分析将10个居群分为3组;主成分分析表明17个植物学形态特征可归为7个主成分因子,累计贡献率达到81.15%,最大程度上反映了所有材料的表型特征。相关分析结果表明,17个形态指标间存在显著的相关性。通过对表型性状与地理信息中海拔的相关性分析结果表明,分蘖数、节数、叶片长、旗叶长、旗叶宽、花序长、第二外颖长、第二内颖长、花序干重、单茎干重与海拔呈极显著负相关。【结论】野生芒自身所具有的遗传物质差异和地理微环境差异导致了野生芒居群内植株在形态特征方面产生了不同程度的变异,从而保持了居群内的高度多样性。海拔对芒植株的生物产量和生殖器官影响很大。
关键词:野生芒;重庆武陵山区;表型变异
【研究意义】芒Andersson),禾本科(Poaceae)芒属(Mscant/wwAndersson)多年生苇状草本,秆高l ̄2m。遍布于我国海拔1800m以下的山地、丘陵和荒坡原野,也分布于朝鲜、日本[1]。芒的生态型较多,分布较广,适应各种逆境及边际土地,地上部分冬季常枯萎,易于收割和储运[2]。随着社会对能源需求的不断增加,寻找可再生的替代能源成为研究的焦点,而生物质能有希望成为全球最有前景的可再生替代能源[3-4]。芒因具有生物量大、抗性强、高纤维素含量、低灰分等特点,而被人们广泛关注,已被欧美列为具有开发潜力的高效能源植物。【前人研究进展】近年来,国内外对芒的研究主要集中在芒的地理分布[5_6]、抗旱性、抗寒性[9]、重金属耐性[1°42]、植物光合特性[13]、核型及细胞学分析[14“5]、遗传多样性及遗传图谱的构建方面。在芒的形态多样性分析方面,卢玉飞[2°]、黄绍英[2|]、肖亮[22]、聂刚[23]和李强[24]对芒属植物的花粉形态、种子表型、表型多样性进行了研究。表型性状多样性体现了遗传的多样性和环境的多样性,而表型性状变异是植物种群的人工驯化和良种选育的基础,也是植物种质资源收集、保存、评价和利用的依据[25]。武陵山区以武陵山脉为中心,平均海拔在1000m左右,地形绵延起伏,地貌变化复杂,具有独特的气候特点[26]。刘琳[27]通过GIS技术定量分析重庆地区植被净第一生产力(NPP)发现,渝东南翼(武陵山区)的植被NPP最高,植被覆盖好,表明该地区植被类型丰富。【本研究切人点】本研究结合科技部基础资源调查专项,以重庆武陵山区5个区县的10个野生芒居群为研究对象,对野生芒的17个外部形态性状进行调查,通过各性状在居群内和居群间的多样性差异分析来揭示重庆武陵山区野生芒资源的多样性程度和分布规律,为芒种质资源的开发利用和保护提供参考。
1材料与方法
1.1供试材料
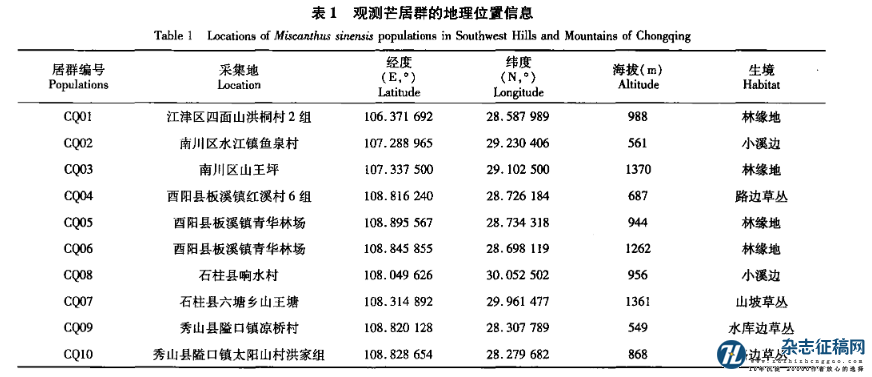
2018年10-11月,对重庆市武陵山区的江津区、南川区、石柱县、酉阳县、秀山县野生芒的分布进行考察。各个区(县)随机选择有代表性的芒居群1 ̄3个,居群间距离3km以上。各居群地理位置和生境信息见表1。
1.2试验内容与方法
表型性状的测量标准参考《牧草种质资源描述规范和数据标准》,每个居群测定20株单株,居群内单株间隔10ra以上。在植株开花期对材料的15个植株形态性状进行测定,包括分蘖数、叶片数、节数、基部茎粗、第一节间长、株高、叶片长、叶片宽、旗叶长、旗叶宽、花序长、第二外颖长、第二内颖长、第二外稃长、基盘毛长,每个单株每项指标重复测定3次取平均值。测定小穗性状时,每株随机选20个小穗测定,重复3次取平均值。随后将材料带回实验室,测定产量性状(单茎干重、花序干重)。测定结果列于表2。
2结果与分析
2.1野生芒居群内和居群间形态特征变异分析
10个芒天然居群性状指标的测量及单因素方差分析结果见表3检验表明,测定的17个数量及质量性状指标在居群间的差异均不显著(P>0.05)。通过F值比较,发现穗部性状指标内颖长在17个性状指标中F值最大(1.12),花序长f值最小(0.29)。

2.2野生芒居群性状的相关分析
对17个表型性状和采集地海拔进行相关分析(表6)。结果表明,野生芒居群表型性状间存在明显的相关性。海拔作为地理信息被引入相关性分析,结果可以看出,海拔与分蘖数、节数、叶片长、旗叶长、旗叶宽、花序长、第二外颖长、第二内颖长、花序干重、单莲干重呈极显著负相关,与盘基毛长呈极显著正相关,与第一节长呈显著正相关。随着海拔增加,温度降低,影响单茎性状的分蘖数、节数、叶片长变小,引起单茎干重下降,同时,旗叶长、旗叶宽、花序长、第二外颖成、第二内颖长这些穗部性状相关指标也递减,最终引起花序干重下降。
3讨论
3.1重庆武陵山区野生芒表型变异的多样性
表型性状研究是植物遗传多样性研究中最基础、最直观的方法,可以从整体上了解研究对象的多样性程度,挖掘更加广泛的基因资源[31]。本研究以重庆武陵山区不同生境下采集的10个野生芒居群为材料,采用Shannon-weaver信息指数对居群的17个表型性状进行遗传多样性分析,得出群体间的平均多样性指数为1.677,居群内平均多样性指数为1.880,因此,野生芒居群的遗传变异主要体现在居群内。居群内的变异主要受遗传作用,居群间的变异受遗传和环境共同作用[32]。Hamrick和Godt等[29]对涉及165个属、499个种的不同类型植物遗传变异和居群分化程度进行统计,结果发现异交风媒植物只有9.9%的遗传变异存在于居群间。芒作为异花授粉、自交不亲和植物[1],其居群内遗传多样性丰富,变异程度大,这与茶条槭[33]和野生玫瑰[34]等植物的表型多样性研究结果相同。笔者在资源调查时发现,同一居群内存在个别植株内稃有芒/无芒及盘基毛为黄色/紫色2种情况,因此,芒自身所具有的遗传物质差异和地理微环境差异导致了野生芒居群内植株在形态特征方面产生了不同程度的变异,从而保持了居群内的高度多样性。
3.2重庆武陵山区野生芒表型地理变异规律
植物形态变异或表型多样性研究,往往采用对形态性状与地理、生态因子进行相关分析以获取植物某些性状与地理或生态因子之间的变化规律[23]。本文对野生芒17个表型性状与海拔进行相关性分析,结果表明,分蘖数、节数、叶片长、旗叶长、旗叶宽、花序长、第二外颖长、第二内颖长、花序干重、单茎干重与海拔呈极显著负相关;随着海拔升高,气温降低,紫外线照射增强,芒植株的分蘖数减少,节数变少,叶片、花序变短,最终导致了花序重量和单茎重量变轻。因此,海拔对芒植株的生物产量和生殖器官影响很大。
参考文献:
[I]陈守良,刘亮,孙必兴.中国植物志[M].北京:科学出版社,1997.
[2]WeijdeTVD,KameiCLA,SeveringEL,etal.Geneticcomplexityofmiscanthuscellwallcompositionandbiomassqualityforbiofuels[J].BMCGenomics,2017,18:406-421.
[3]LewandowskiI,ScurlockJM0,LindvallEl,etal.ThedevelopmentandcurrentstatusofperenmalliiizomatousgrassesasenergycropsintheUSandEurope[J].BiomassandBioenei^y,2003,25:335—361.
[4]梁绪振,陈太祥,白史且,等.芒属(施ca/U/^s)植物种质资源研究进展[J].草业与畜牧,2010(10):1-5.
[5]肖亮,易自力.四种芒属植物的地理分布及气候模式研究[J]?草地学报,2017,25(4):685-690.
何玮,冉启凡,徐远东,张丽、范彦
相关阅读
论文常识
期刊知识
著作出版
教材出书
专利申请
出版社
 扫码关注公众号
扫码关注公众号